16.7 – 15.8 million years
Horses have probably been the single most important domesticated animal in human history. Also, more than with other livestock, people get attached to horses as individuals. I’m guessing that in history and literature there are more horses with individual names than any other animal. (Alexander the Great’s horse was Bucephalus, “Ox-head”; Muhammed’s was al-Buraq*; Charlemagne’s was Tencendur; Don Quixote’s was Rocinante; Gandalf’s was Shadowfax.) We’ll be hearing a lot more about horses and horse folk on Logarithmic History once we get to human history.
Being so charismatic, horses have featured in a big way in arguments over evolution. Thomas Henry Huxley (1825-1895), “Darwin’s bulldog,” knew he needed to find good evidence for evolution. When he visited the United States in 1876, he was ready to give a lecture based on horse fossils from Europe. But visiting Yale, he was so impressed with O. C. Marsh’s collection of horse fossils from the western United States, that he rewrote his lecture around it.
Henry Fairfield Osborn (1857-1935) was director of the American Museum of Natural History and a huge presence in American paleontology. He was active at a time when most scientists accepted evolution, but many weren’t so keen on Darwin’s theory of natural selection. He thought horses were a fine example of “orthogenesis,” the tendency of species to follow a fixed line of evolution, reflecting internal forces, maybe related to willpower. He thought that humans shared a migratory spirit with horses, so that anywhere horse fossils were found would be a good place to look for human fossils. This theory didn’t pan out too well. A massive AMNH expedition to Central Asia led by Ray Chapman Andrews found all sorts of wonders – dinosaur eggs, baluchitheres – but no fossil “pro-men.” Orthogenesis leant itself naturally to diagrams showing evolution from early to modern horses going in a straight line.

George Gaylord Simpson (1902-1984), paleontologist, was one of the great figures in the evolutionary Modern Synthesis that brought together Darwin’s theory of natural selection and Mendel’s genetics. There was no room for orthogenesis in the Modern Synthesis, and Simpson emphasized that the evolution of horses was a matter of adaptation to a changing environment – especially the spread of grasslands. Also that horse evolution looked more like a bush than a ladder.

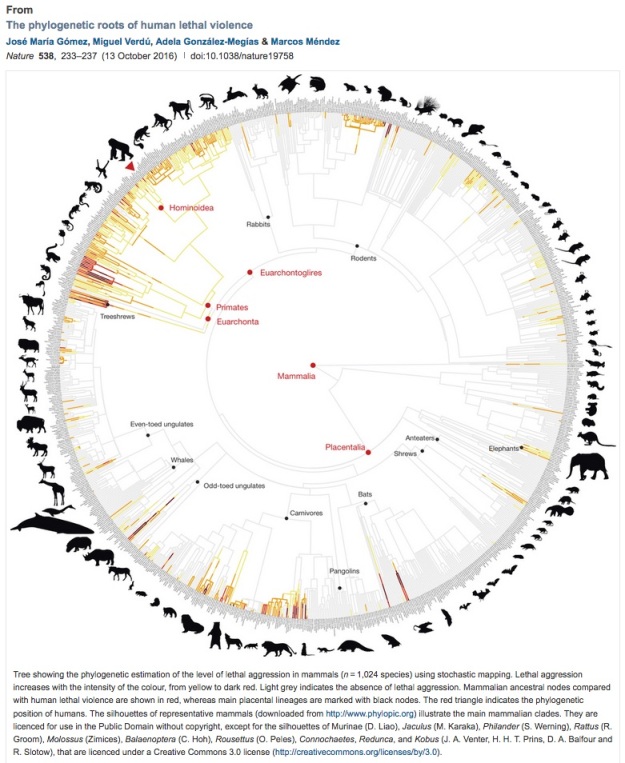
Stephen Jay Gould (1941-2002) was the most widely recognized American evolutionary biologist of recent times. (For example had a spot on The Simpson’s — “Lisa The Skeptic,” Season 9.) Gould had his own take on the modern synthesis, taking the “bushes not ladders” theme for horses and other animals (including human ancestors), and pushing it a step further. According to the theory of “punctuated equilibrium” (formulated in collaboration with Niles Eldredge), species mostly change relatively little during the time they exist (evolutionary stasis). Most evolutionary change happens when a small population buds off to form a new species and reproductive isolation allows it to conserve any evolutionary novelties it has developed. This opens up the possibility of “species selection.” Applied to horses, for example, this could mean that horses were evolutionarily successful for some time not so much because individual horses were well-adapted, but because something about horses collectively (their harem mating system, maybe) made one horse species especially likely to generate new species. Both horses and primates seem to be especially prone to bud off new species:
Speciation and chromosomal evolution seem fastest in those genera with species organized into clans or harems (e.g., some primates and horses) or with limited adult vagility and juvenile dispersal, patchy distribution, and strong individual territoriality (e.g., some rodents). This is consistent with the … hypothesis … that population subdivision into small demes promotes both rapid speciation and evolutionary changes in gene arrangement by inbreeding and drift.
* Richard Dawkins doesn’t believe that Muhammed’s horse, al-Buraq, carried him (i.e. Muhammed) to heaven and back.